INTRODUCTION
Periprosthetic osteolysis is a common complication after total ankle arthroplasty (TAA), in which numerous types of cells communicate through an intricate paracrine network of cytokines, chemokines, and other molecules, resulting in bone loss or loosening of implant.[1] Although TAA has achieved promising clinical results in the treatment of end-stage ankle arthritis, aseptic loosening is a common cause for revision TAA surgery.[2] Aseptic loosening and periprosthetic osteolysis occur due to the biological response to particulate wear debris. Particle-stimulated macrophages and other cells release cytokines, chemokines, and other pro-inflammatory substances that perpetuate chronic inflammation, eventually leading to local bone loss and decreased new bone formation.[1,3] Osteolysis is a complex condition that results from the production of several macrophage-derived cytokines, such as tumor necrosis factor (TNF), interleukin (IL)-1, and IL-6, in response to implant-derived debris at the site of bone implants, leading to osteoclast activation and bone resorption at the implant interface.[4] In a titanium particle-induced osteolysis model, the level of inflammatory factors was significantly increased, promoting osteoclast differentiation.[5] Although osteoblasts may not be the main players among macrophages, they play an important synergistic role by cooperating with the macrophages and osteoclasts during osteolysis. In addition, implant-derived debris in periprosthetic osteolysis may affect osteoblast maturation and function, thereby contributing to osteolysis by decreasing bone formation.[6,7]
Bone metabolism equilibrium is maintained by osteoclasts via bone resorption function in cooperation with osteoblasts via bone formation function. Functional disorders of osteoclasts and osteoblasts, particularly an abnormal increase in osteoclast activity, are responsible for several diseases, such as osteoporosis, periprosthetic osteolysis, bone tumors, and Paget’s disease.[1,8,9] Since receptor activator of nuclear factor-κB ligand (RANKL)/ receptor activator of nuclear factor-κB (RANK) signaling regulates the formation of multinucleated osteoclasts from their precursors, as well as their activation and survival in normal bone remodeling and various pathologic conditions, recent studies have shown that the RANKL/RANK/osteoprotegerin (OPG) pathway plays an important role in osteoclastogenesis, as well as in osteolysis.[1,10]
LIM homeobox transcription factor 1-β (Lmx1b), a LIM-homeodomain family of transcription factors, plays an important role in the normal development of multiple tissues, such as the development of the limb, brain, spinal cord, kidney, and eye tissues.[11,12] Cesario et al. [13] reported that Lmx1b plays an important role in specifying the potential of different regions within the head mesenchyme with different osteogenic potentials at the beginning of calvarial bone development but not during limb development. In addition, we recently reported that Lmx1b acts as a negative regulator during osteoblast differentiation and bone formation by regulating Runt-related transcription factor 2 (Runx2) activity.[14] However, despite research on the function of Lmx1b in development and differentiation of osteoblast, the role of Lmx1b in osteoclasts is yet to be elucidated.
In this study, we aimed to identify a molecular biomarker for osteolysis after TAA and investigate the role of the identified genes in osteoclast differentiation. We found that Lmx1b expression was increased in patients with periprosthetic osteolysis after TAA. Through gain-of-function experiments, we demonstrated that Lmx1b acts as a negative regulator in RANKL-induced osteoclast differentiation via the regulation of nuclear factor of activated T cells 1 (NFATc1).
METHODS
1. RNA sequencing analysis
For RNA sequencing, eight patient samples were collected: 5 primary TAA samples (control group) and 3 TAA samples revised for flexion instability (osteolysis group). The study was approved by the Institutional Review Board (IRB) of Chonnam National University Hospital (IRB no. BTMP-2019-055). All patients were identified, and oral and written informed consent with signatures were obtained prior to enrollment. At the time of revision surgery, the suprapatellar synovial capsular tissues (either control or osteolysis group) were carefully dissected by the operative surgeon using a scalpel. After rinsing with phosphate buffered saline (PBS), the tissues were frozen in liquid nitrogen. Total RNA was extracted using the RNeasy Kit (Qiagen GmbH, Hilden, Germany), according to the manufacturer’s protocol. RNA sequencing and data analysis were performed by Ebiogen Inc. (Seoul, Korea). The RNA quality was determined using an Agilent 2100 bioanalyzer system (Agilent Technologies Netherlands B.V., Amstelveen, the Netherland). Library construction was performed using the Quant-Seq library prep kit (Lexogen, Vienna, Austria), according to the manufacturer’s protocol. High-throughput sequencing was performed using Illumina HiSeq 2000 (Illumina, San Diego, CA, USA). The analysis of differentially expressed genes and gene ontology was performed using the Excel-based Differentially Expressed Gene Analysis (ExDEGA) software package (Ebiogen Inc.).
2. Osteoclast differentiation, tartrate-resistant acid phosphatase (TRAP) staining, and pit formation analysis
Cell culture media and supplements were obtained from HyClone (HyClone Laboratories, Lagan, UT, USA). Recombinant human macrophage colony-stimulating factor (M-CSF) and recombinant human sRANKL were purified from Escherichia coli. Murine osteoclasts were prepared from bone marrow cells obtained by flushing femurs and tibiae from 6 week-old ICR mice. Bone marrow cells were cultured in α-minimal essential medium containing 10% fetal bovine serum (FBS) with M-CSF (30 ng/mL) for 3 days, and attached cells were used as osteoclast precursor (bone marrow-derived macrophages [BMMs]). To generate osteoclasts, BMMs were cultured with M-CSF (30 ng/mL) and RANKL (100 ng/mL) for 3 days. All cells were cultured at 37°C and 5% CO2. Cultured cells were fixed and stained with TRAP. TRAP-positive multinuclear cells, containing more than 3 nuclei, were counted as osteoclast. Cells were observed using a Leica DMIRB microscope equipped with an N plan 10×0.25 numerical aperture objective lens (Leica Microsystems, Wetzlar, Germany). Images were obtained with a ProgRes CFscan (Jenoptik, Jena, Germany) camera and using a ProgRes Capture Pro (Jenoptik). For pit formation analysis, BMMs were cultured with M-CSF and RANKL on Osteo assay plates (Corning Inc., Corning, NY, USA) for 4 days. Resorption lacunae were visualized by bright-field light using a Leica DMIRB microscope with a ProgRes CFscan (Jenoptik) camera and using a ProgRes Capture Pro (Jenoptik).
3. Quantitative real-time polymerase chain reaction (PCR) analysis
Cells were lysed in QIAzol (Qiagen GmbH), and total RNA was isolated from the cultured cells according to the manufacturer’s protocol. Purified RNA was reverse transcribed into cDNA using GoScriptTM Reverse Transcriptase (Promega, Madison, WI, USA). To examine the mRNA expression levels, the cDNA synthesized was used for SYBR-based real-time PCR performed in triplicate using a Rotor-Gene6 instrument (Qiagen GmbH). The thermal cycling conditions were as follows: 15 min at 95°C, followed by 40 cycles at 94°C for 15 sec, 55°C for 30 sec, and 72°C for 30 sec. The mRNA concentrations were normalized to an endogenous housekeeping gene, glyceraldehyde 3-phosphate dehydrogenase (GAPDH). The relative quantitation value for each target gene compared with the calibrator for the target was expressed as 2−(Ct-Cc) (Ct and Cc are the mean threshold cycle differences after normalizing to GAPDH). The relative expression levels of the samples were presented as a semi-log plot. The primer sequences are listed in Table 1.
4. Retroviral gene transduction
Plat-E cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM; HyClone Laboratories, Lagan, UT, USA) supplemented with 10% FBS. To prepare retroviral supernatants, recombinant plasmids and the parental pMX vector were transfected into the packaging cell line Plat-E using Fugene6 (Promega) according to the manufacturer’s protocol. Viral supernatant was collected from the cultured media 48 hr after transfection. Osteoclast precursor cells or osteoblast precursor cells were incubated with the viral supernatants for 6 hr in the presence of 10 μg/mL polybrene (Sigma-Aldrich, St. Louis, MO, USA).
5. Immunoprecipitation and fractionation
For immunoprecipitation, 293T cells transfected with Flag-Lmx1b and hemagglutinin (HA)-NFATc1 were harvested after washing with ice-cold PBS and then lysed in the extraction buffer (50 mM Tris-HCl, pH 8.0, 150 mM NaCl, 1 mM EDTA, 0.5% Nonidet P-40, and 0.01% protease inhibitor cocktail). Lysates were immunoprecipitated with their corresponding antibodies. Cell lysates and precipitated samples were subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene fluoride membranes (Millipore, Billerica, MA, USA). For nuclear fractionation, the cultured cells were harvested after washing with prechilled PBS, and the cells were fractionated using NE-PER Nuclear and cytoplasmic extraction reagent (Thermo Fisher Scientific, lnc., Waltham, MA, USA), according to the manufacturer’s protocol. Cytoplasmic and nuclear extracts were subjected to SDS-PAGE and western blotting. The primary antibodies include NFATc1, LaminB1 (Santa Cruz Biotechnology, Dallas, TX, USA), HA, and Flag (Sigma-Aldrich). HRP-conjugated secondary antibodies (Abcam, Cambridge, MA, USA) were probed and developed with ECL solution (Millipore). Signals were detected and analyzed using the Azure c300 Chemiluminescent Western Blot Imaging System (Azure Biosystems Inc., Dublin, CA, USA).
6. Immunohistochemistry
For the immunohistochemical staining of Lmx1b, paraffin-embedded tissues were cut into 5-μm thick serial slices. The sections were deparaffinized in xylene at room temperature for 20 min, followed by rehydration through a graded series of alcohol solutions. Following the peroxidase blocking of the tissue slides, the slides were incubated overnight with primary antibody against Lmx1b (Abcam) at a dilution of 1:100 at 4°C, and hematoxylin was used as a counterstain.
RESULTS
1. Profile of differentially expressed genes in patients who underwent revision TAA
To identify molecular biomarkers for osteolysis after TAA, RNA-seq was performed using tissue biopsy in patients who underwent revision implant surgery due to peri-implant osteolysis. Details of the patients in each group are presented in Figure 1A. The mean age was 64.3±3 years and 68.7±5 years in the control and osteolysis groups, respectively, and the mean body mass index was 23.6±1.8 kg/m2 and 23.9±2.3 kg/m2 in the control and osteolysis groups, respectively. A difference of over 2-fold relative to the control was considered significant. We found that 1,235 genes were significantly dysregulated in the osteolysis group compared to that in the control group: 736 genes were upregulated and 499 genes were downregulated in the osteolysis group. Analyses of differentially expressed genes and gene ontologies indicated that the expression of LMX1B, doublecortin (DCX), trefoil factor 3 (TFF3), pro-platelet basic protein (PPBP), ring finger protein 165 (RNF165), KIAA0319, opioid receptor mu 1 (OPRM1), transforming growth factor-β1 (TGFB1), TNF ligand superfamily member 11 (TNFSF11; RANKL), and p21-activated kinase 1 (PAK1) was upregulated, while the expression of FERM domain containing kindlin 1 (FERMT1), POU class 2 homeobox 3 (POU2F3), leukotriene B4 receptor 2 (LTB4R2), von Willebrand factor A domain containing 2 (VWA2), TNF, IL1, IL6, and TNF receptor superfamily member 11b (TNFRSF11B; OPG) was downregulated in the osteolysis group; no difference was observed in the expression of TNF receptor superfamily member 11a (TNFRSF11A; RANK) (Fig. 1B). Next, the relative mRNA expression levels of these genes in patients with osteolysis following TAA were examined in tissues obtained from patients who underwent a revision TAA using real-time PCR. Among the molecules examined, the mRNA expression of LMX1B, DCX, TFF3, and PPBP was found to be significantly increased in the osteolysis group. However, no difference was observed in the mRNA expression of RNF165, OPRM1, PAK1, VWA2, FERMT1, LTB4R2, and POU2F3 between the control and osteolysis group, while the mRNA expression of KIAA0139 was decreased in the osteolysis group (Fig. 1C).
2. Lmx1b inhibits RANKL-induced osteoclast differentiation
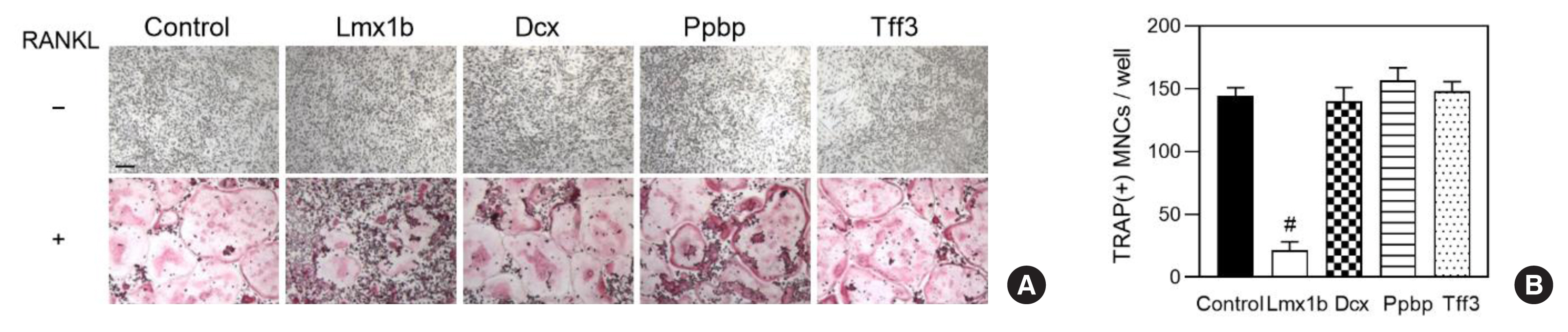
Since the expression of some genes was increased in the osteolysis group, the role of these genes in osteoclast differentiation was examined next. To this end, retroviral vectors containing the genes with an increased expression in the osteolysis group (Fig. 1) were constructed. The resulting control or retroviral vectors were then overexpressed in osteoclast precursor cells via retroviral transduction. The primary osteoclast precursor cells were infected with the control or retroviral vectors containing the genes with an increased expression in the osteolysis group and further cultured with M-CSF and RANKL. The number of TRAP-positive multinucleated cells was increased in the control vector-infected cells. Contrary to our expectations, Lmx1b overexpressing osteoclasts exhibited a strong inhibition of RANKL-induced osteoclast formation, whereas the effect of other genes, Dcx, Ppbp, and Tff3, did not differ from that of the control (Fig. 2A, B). Therefore, the subsequent analysis focused on Lmx1b in osteoclast differentiation.
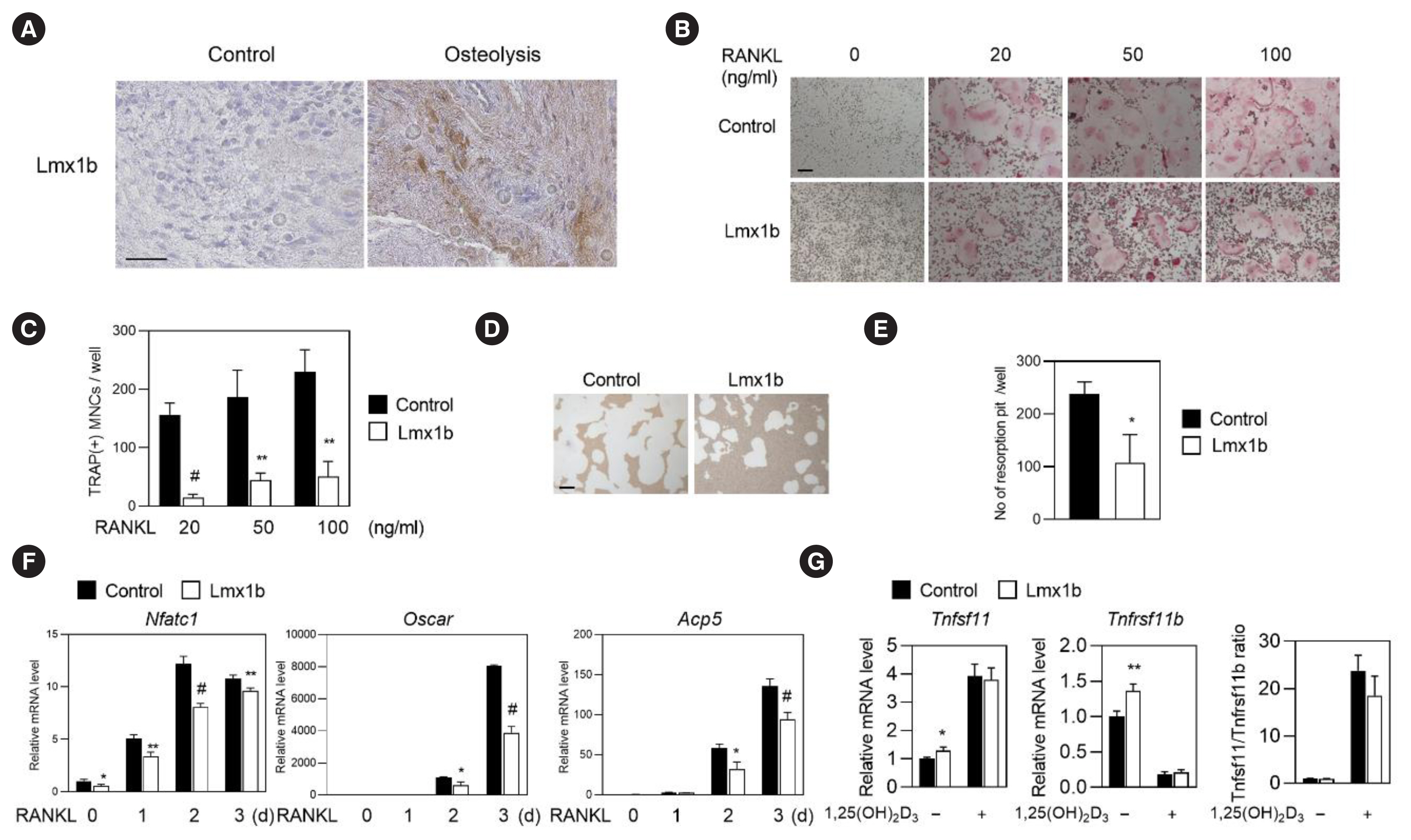
3. Expression of Lmx1b is upregulated in the osteolysis group and Lmx1b overexpression inhibits RANKL-induced osteoclast differentiation
To evaluate the expression of Lmx1b in the osteolysis group, paraffin sections of tissues from patients who underwent revision arthroplasty for periprosthetic osteolysis were evaluated using immunohistochemistry. The results showed that Lmx1b was highly expressed in the tissues obtained from patients with osteolysis compared to that in the tissues from healthy individuals (Fig. 3A). As the expression of Lmx1b was higher in the osteolysis group than in the control group and Lmx1b overexpression inhibited osteoclast formation, the role of Lmx1b during osteoclast differentiation was evaluated further. To this end, primary osteoclast precursors were infected with the control or Lmx1b retrovirus and cultured with various concentration levels of RANKL. Lmx1b overexpression was found to significantly attenuate osteoclast formation at the various concentration levels of RANKL (Fig. 3B, C). Next, when Lmx1b overexpressed precursors were cultured on the surface of a synthetic calcium phosphate matrix in the presence of M-CSF and RANKL, the formation of resorption pits was significantly reduced (Fig. 3D, E). In addition, Lmx1b overexpression strongly attenuated the RANKL-mediated induction of Nfatc1, osteoclast-associated receptor (Oscar), and acid phosphatase 5 (Acp5) (Fig. 3F).
Osteoblasts regulate the expression of various genes, especially RANKL and OPG, thereby contributing to osteoclast activation during normal bone remodeling, as well as in a variety of pathological conditions, including osteolysis.[1,15] The 1,25-dihydroxy-cholecalciferol (1,25[OH]2D3) indirectly stimulates osteoclast differentiation by increasing the expression of Tnfsf11 (RANKL) and decreasing the expression of Tnfrsf11b (OPG) in osteoblasts in vitro.[16] Thus, whether Lmx1b plays a role in the 1,25(OH)2D3-mediated induction of Tnfsf11/Tnfrsf11b ratio in osteoblasts was investigated. However, Lmx1b overexpression was not found to have a significant effect on Tnfsf11 and Tnfrsf11b expression (Fig. 3G). Collectively, these results suggest that Lmx1b negatively regulates RANKL-induced osteoclast differentiation without affecting of the 1,25(OH)2D3 dependent Tnfsf11 and Tnfrsf11b expression in osteoblast.
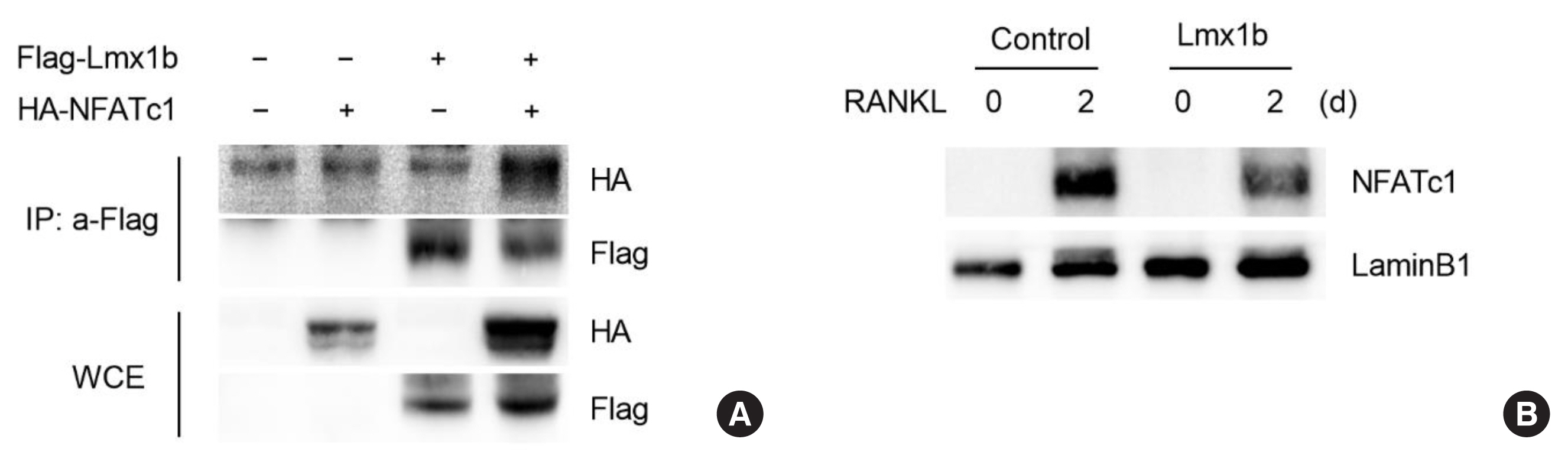
4. Lmx1b regulates NFATc1 activity
RANKL stimulation increases the expression of NFATc1, which is a key transcription factor during osteoclast differentiation. The induction of NFATc1 is followed by the induction of OSCAR and TRAP, which are osteoclast-specific genes.[17] To further investigate the molecular mechanism by which Lmx1b regulates NFATc1 expression in RANKL-induced osteoclast differentiation, whether Lmx1b interacts with NFATc1 was examined. To this end, 293T cells were transfected with HA-NFATc1 or Flag-Lmx1b and a co-immunoprecipitation assay was performed. As shown in Figure 4A, an interaction was observed between Lmx1b and NFATc1. RANKL is known to activate calcium/calmodulin-dependent calcineurin pathways in the cytoplasm, followed by NFATc1 nuclear translocation, as well as activate NFATc1 to induce differentiation of osteoclast.[18] Therefore, the effect of Lmx1b on nuclear translocation of NFATc1 was evaluated, and the overexpression of Lmx1b was found to suppress NFATc1 nuclear translocation compared to the control (Fig. 4B). Taken together, these results suggest that Lmx1b negatively regulates osteoclast differentiation by blocking the nuclear localization of the NFATc1 protein.
DISCUSSION
Although TAA has achieved promising clinical results in the treatment of end-stage ankle arthritis and periprosthetic osteolysis, subsequent aseptic loosening is a common cause for revision surgery. Many studies have been conducted to establish the function of osteoclasts, which can resorb bone in aseptic loosening.[19,20] In this study, RNA-seq analysis using tissues from patients who underwent revision arthroplasty was performed to identify osteolysis-related molecular biomarkers of periarthroplasty osteolysis after TAA. Using real-time PCR and immunohistochemistry analyses, we found that Lmx1b expression was upregulated in the periarthroplasty osteolysis group after TAA. These results suggest that upregulated Lmx1b may modulate osteolysis by enhancing osteoclast differentiation. However, unexpectedly, Lmx1b overexpression in osteoclast precursor cells attenuated RANKL-induced osteoclast differentiation (Fig. 3).
We recently reported that Lmx1b acts as a negative regulator of Runx2 in BMP2 signaling pathway in osteoblast and bone regeneration.[14] Since Lmx1b was more abundantly expressed in osteoblasts than in osteoclasts, it can be deduced that, although Lmx1b negatively regulates differentiation of both osteoclasts and osteoblasts, the net effect of overexpression of Lmx1b may be stronger in osteoblasts than that of Lmx1b in osteoclasts during periprosthetic osteolysis after TAA. This highlights the possibility that upregulated Lmx1b expression in osteoblasts at the osteolysis site is involved in the decrease of bone matrix, which results in reduced bone mass. However, further study on the regularity effect of increased Lmx1b expression in periprosthetic osteolysis is needed to elucidate the complex mechanisms underlying TAA-induced bone loss.
Several studies have shown that various factors, including IL-1, IL-6, IL-8, monocyte chemotactic protein-1, vascular endothelial growth factor, RANKL, prostaglandin E2, OPG, TNF, and TGF-β, are present in the periprosthetic region.[4,21-23] These soluble factors can be easily delivered to the bone cells and have instantaneous effects. Using RNA-seq analysis, the expression of TGF-β and RANKL was upregulated, while the expression of OPG was downregulated in the periarthroplasty osteolysis group. However, using differentially expressed gene analysis, the expression of cytokines with a well-known function in bone resorption, such as IL-1, TNF, and IL-6, was found to unexpectedly decrease in the osteolysis group (Fig. 1B). This suggests that various patient characteristics, including age, weight, underlying disease, and activity level, in addition to the operative materials, can affect the amount and type of osteolysis.
Although research on osteolysis is primarily based on osteoclasts, macrophages, and fibroblasts, osteoblasts are also known to participate in bone degeneration by altering the expression of chemokines and proteinase, which act in a paracrine manner to recruit inflammatory cells into the periprosthetic space and to stimulate osteoclastic bone resorption. For example, increased TNFα secretion is known to induce the release of IL-6, IL-8, monocyte chemoattractant protein-1, TGF-β, and RANKL in osteoblasts, while the release of IL-6 may indirectly activate osteoclasts by upregulating RANKL expression in the periprosthetic space.[4,16, 24-26] 1,25(OH)2D3 indirectly stimulates osteoclast differentiation by increasing the expression of Tnfsf11 (RANKL) and decreasing the expression of Tnfrsf11b (OPG) in osteoblasts.[15] Among the aforementioned cytokines, the Tnfsf11/ Tnfrsf11b ratio is an important determinant of bone mass in both normal and disease conditions. Thus, whether Lmx1b affects 1,25(OH)2D3-mediated Tnfsf11/Tnfrsf11b ratio in osteoblasts was investigated. However, overexpressed Lmx1b in osteoblasts was not found to affect the Tnfsf11/Tnfrsf11b ratio (Fig. 3). Collectively, these results highlight the possibility that Lmx1b exerts its action in osteoclast differentiation without affecting the Tnfsf11/Tnfrsf11b ratio in periprosthetic osteolysis.
In this study, Lmx1b overexpression was demonstrated to inhibit osteoclast differentiation. Lmx1b was found to inhibit NFATc1 activity in osteoclasts by blocking the nuclear translocation of NFATc1. The study on Lmx1b in osteoclasts can be applied to a range of diseases, including osteoporosis, caused by excessive osteoclast activity. The results presented in this study provide a promising basis for the development of therapeutic strategies responsible for the prevention and treatment of periprosthetic bone loss.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print