Bone Morphogenetic Protein-2 Desensitizes MC3T3-E1 Osteoblastic Cells to Estrogen Through Transcriptional Downregulation of Estrogen Receptor 1
Article information
Abstract
Background
Estrogens exert preferable effects on bone metabolism through two estrogen receptors (ERs), ER1 and ER2, which activate the transcription of a set of genes as ligand-dependent transcription factors. Thus, growth factors and hormones which modulate ER expression in the bone, if any, may possibly modulate the effect of estrogens on bone metabolism. However, research as to which of these molecules regulate the expression of ERs in osteoblasts has not been well documented.
Methods
A reporter assay system developed in this study was used to explore molecules that modulate ER1 expression in MC3T3-E1 osteoblastic cells. Gene expression was analyzed by reverse transcription-polymerase chain reaction.
Results
A pilot study using the reporter system revealed that bone morphogenetic protein (BMP)-2 negatively regulated ER1, but not ER2, expression in MC3T3-E1 cells. Consistently, estradiol-induced reporter activity via an estrogen responsive element was strongly suppressed in MC3T3-E1 cells pretreated with BMP-2.
Conclusions
BMP-2 desensitizes osteoblastic cells to estrogen through downregulation of ER1 expression.
INTRODUCTION
Estrogen plays an important role in various tissues, including the bone, brain and heart. In the bone of women and female animals, estrogen exerts a preferable effect in its turnover.[1,2] Conversely, estrogen deficiency in postmenopausal women engenders accelerated bone resorption thereby leading to osteoporosis.[3] The actions of estrogen are known to be mediated by two isoforms of nuclear receptors, estrogen receptor (ER)1 and ER2 (also designated as ERα and ERβ, respectively), which also function as ligand-dependent transcriptional factors. In the bone, both osteoblasts (bone-forming cells) and osteoclasts (bone-resorbing cells) have been demonstrated to express ERs,[4] indicating that estrogen directly acts on bone turnover. The two ER isoforms exhibit different properties in terms of ligand-specificity, tissue distribution and downstream intracellular signaling.[5-7] These differences were reflected by the distinct phenotypes of mice, which were deficient in either or both of ER1 and ER2.[8-10] To date, the interplay between estrogen and bone-related growth factors, e.g., transforming growth factor (TGF)-β[11-13] and bone morphogenetic proteins (BMPs),[14] in osteoblastic cells has been demonstrated. Further, the signaling cross-talk between estrogen and interleukin-6 has also been reported.[15] However, research on how the expression of ERs in osteoblasts are regulated by bone-related growth factors or hormones has not been well documented.
In order to search for bioactive molecules that regulate the expression of ERs in osteoblasts at the transcriptional level, this study developed MC3T3-E1 osteoblastic cell lines which are stably transfected with a reporter construct that has the luciferase gene under the control of a mouse ER1 gene promoter. By conducting a pilot study using this reporter system, the research also identified BMP-2 as a possible suppressor of ER1 expression. Through a reverse transcription-polymerase chain reaction (RT-PCR), the study also verified that mRNA expression of ER1, but not that of ER2, in MC3T3-E1 was significantly downregulated by BMP-2. Further, the research also demonstrated that the responsiveness of MC3T3-E1 cells to estradiol (E2) was pronouncedly suppressed by the pretreatment with BMP-2. These results indicate that BMP-2 is a possible negative regulator of ER1 gene expression in osteoblastic cells.
METHODS
1. Reagents
Recombinant human BMP-2 (rhBMP-2) and 1alpha,25-dihydroxy-vitamin D3 (1,25-[OH]2D3) were kindly provided by Astellas Pharma Co. Ltd (Tokyo, Japan). E2 and recombinant mouse epidermal growth factor (rmEGF) were purchased from Sigma-Aldrich Co. (St. Louis, MO). Phorbol-12-myristate-13-acetate (PMA) and dibutyryl cyclic adenosine monophosphate (dbcAMP) were purchased from Wako (Osaka, Japan).
2. Cell culture
Mouse calvaria-derived MC3T3-E1 osteoblastic cells were purchased from Riken Bioresource Center (Tsukuba, Ibaraki, Japan). The cells were maintained in α-minimum essential medium (Invitrogen, Carlsbad, CA, USA) containing 10% fetal bovine serum at 37℃ in an atmosphere of 5% CO2/95% air.
3. Construction of reporter plasmids
A mouse genomic DNA fragment encompassing the ER1 promoter region of 2.8 kbp upstream of its transcription initiation site was obtained by screening of a mouse genomic DNA library using the 32P-labeled 5'-untranslated region of a mouse ER1 cDNA as a probe. The fragment was then subcloned into a pGVB2 vector (Nippongene, Tokyo, Japan) at its multicloning site. A fragment encompassing a neomycin-resistance gene derived from pSV2neo was then inserted into the vector in order to construct a final plasmid designated pMERAluc1 (Fig. 1).
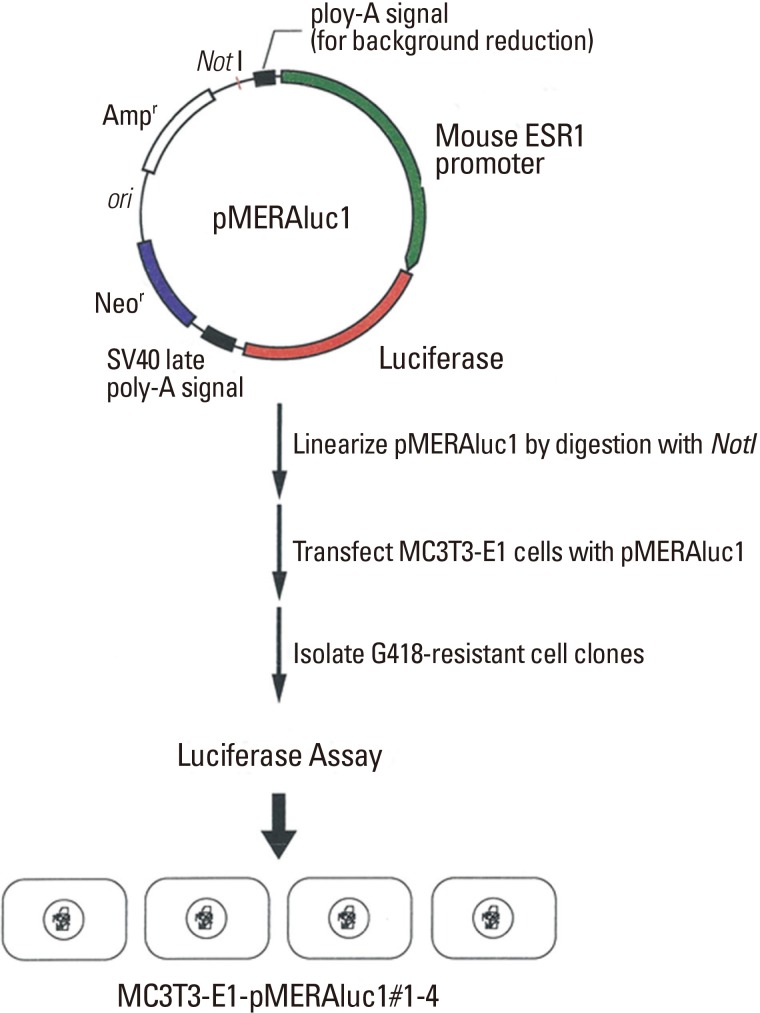
Construction of the reporter plasmids used in this study.
4. Stable transfection of MC3T3-E1 cells with the reporter construct under control of ER1 gene promoter
To establish a reporter system for identifying bioactive molecules in order to modulate ER1 gene expression, MC3T3-E1 cells were transfected with pMERAluc1. MC3T3-E1 cells were transfected with linearized pMERAluc1 using FuGENE 6 transfection reagent (Roche Diagnostics, Basel, Switzerland) and cultured in the presence of 500 µg/mL G418 (Invitrogen) for 2 weeks. G418-resistant cells which formed colonies were separately trypsinized using metal cloning rings and transferred to wells of 24-well culture plates to further culture in the presence of 500 µg/mL G418. When reaching semi-confluence, these cells were expanded in a larger culture apparatus to allow propagation; finally, 4 clones of these cells (designated MC3T3-E1-MERAluc1#1-4) could be successfully isolated as stably transformed cell lines.
5. Luciferase reporter assay
ER1 promoter-driven transcription of the luciferase gene in MC3T3-E1 cells was evaluated by luciferase reporter assay using Picagene Luminescence Kit (Wako). The cells were lysed in LCβ lysis buffer (Wako), and luciferase activities in the cell lysates were measured using a TD-20/20 luminometer (Promega, Madison, WI, USA), as previously described.[16]
6. Transient transfection
To validate the results obtained using the stable clones MC3T3-E1-MERAluc1#1-4, MC3T3-E1 cells were also transfected transiently with pMERAluc1. Transfection of the cells were performed using a FuGene6 transfection reagent, as previously described.[7] Twenty-four hours after the transfection, rhBMP-2 was added at a final concentration of 250 ng/mL. After 24-hour exposure with rhBMP-2, the cells were harvested and lysed in LCβ lysis buffer. The measurement of luciferase activity in the lysates were performed as described above.
7. RT-PCR
Cellular total RNAs were extracted using TRIzol reagent (Invitrogen, Carlsbad, CA, USA), according to the manufacturer's protocol. RT-PCR for mouse ER1, ER2 and glyceraldehyde-3-phosphate dehydrogenase gene (GAPDH) was performed as described earlier [16]. Primers used in this study are listed in Table 1.

Primers used in this study
8. Statistical analysis
Statistical analysis was performed using an unpaired student's t test. The results are shown as the means±SD.
RESULTS
1. Establishment of MC3T3-E1 cell lines stably harboring ER1 gene promoter-driven luciferase reporter
By conducting the procedures described in the Materials and Methods section, four discrete MC3T3-E1 cell clones stably harboring the luciferase reporter gene downstream of ER1 gene promoter, which were designated as MC3T3-E1-MERAluc1#1-4 (Fig. 1), were finally isolated.
2. Identification of factors regulating ER1 mRNA expression in MC3T3-E1 cells
Relative luciferase activities in MC3T3-E1-MERAluc1 cells treated with several bioactive molecules, such as 1,25-(OH)2D3, E2, 4-hydroxy-tamoxifen (4-OH-Tam), PMA, dbcAMP, rmEGF and rhBMP-2, were measured. Of these molecules, 1,25-(OH)2D3, PMA, mEGF and rhBMP-2 decreased the reporter activity driven by the ER1 promoter in all the stable cell clones (Fig. 2A). By contrast, E2, 4OH-Tam and dbcAMP had no effect on the promoter activity (Fig. 2A). In this study, the BMP-2-elicited downregulation of ER1 promoter activity was focused, because the functional interplay between BMPs and estrogens in osteoblasts has not been much addressed so far, although both are well known to play important roles in osteoblast function.
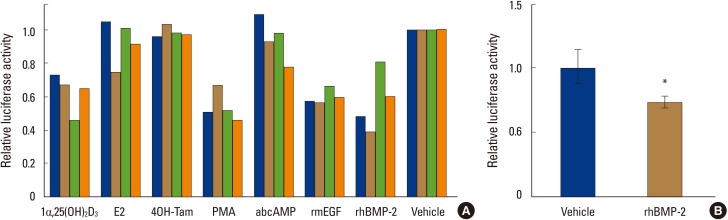
Bone morphogenetic protein-2 downregulates a reporter activity driven by mouse estrogen receptor1 gene promoter in MC3T3-E1 cells. (A) MC3T3-E1-MERAluc1 cells (clone#1-4) were treated with 1alpha,25-dihydroxy-vitamin D3 (1,25-[OH]2D3; 10 nM), estradiol (E2; 100 nM), 4-hydroxy-tamoxifen (4-OH-Tam; 100 nM), phorbol-12-myristate-13-acetate (PMA; 10 nM), dibutyryl cyclic adenosine monophosphate (dbcAMP; 1 mM), recombinant mouse epidermal growth factor (rmEGF; 100 ng/mL) and recombinant human bone morphogenetic protein-2 (rhBMP-2; 250 ng/mL). After a 48-hour incubation, these cells were harvested and lysed then, luciferase activities in the lysates were measured. Luciferase activities in the vehicle-treated cells were set at 1. (B) MC3T3-E1 cells were transiently transfected with pMERAluc1 (0.5 µg) along with pSV-β-gal (0.3 µg) for normalization of transfection efficiency, incubated for 24 hours and exposed by rhBMP-2 (250 ng/mL) or vehicle. Forty-eight hours later, the cells were harvested and lysed in LCβ lysis buffer. Luciferase activities were measured as described in the Materials and Methods section. The results are indicated as relative values when the normalized luciferase activity of the vehicle-treated cells is set at 1. *P<0.05.
To verify the above result, the effect of rhBMP-2 on ER1 gene promoter activity was examined using MC3T3-E1 cells transiently transfected with pMERAluc1. As expected, rhBMP-2 significantly suppressed the ER1 gene promoter-driven luciferase activity in MC3T3-E1 in this experiment (Fig. 2B).
3. BMP-2 suppresses ER1 mRNA expression in MC3T3-E1 cells
The study then examined whether the ER1 gene promoter activity suppressed by BMP-2 was reflected by the alteration in ER1 mRNA expression in BMP-2-treated MC3T3-E1 cells. RT-PCR analyses revealed that ER1 mRNA expression in the cells was suppressed starting from 6 hours after the BMP-2 treatment (Fig. 3A). Conversely, rhBMP-2 did not change the mRNA expression of ER2 (Fig. 3B).
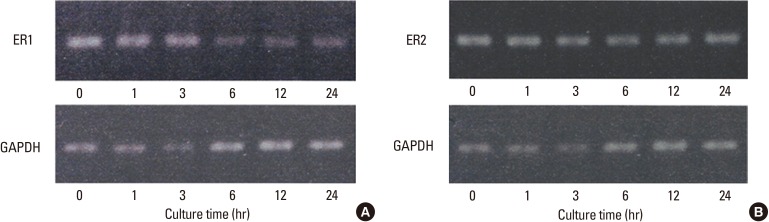
Bone morphogenetic protein-2 downregulates expression of estrogen receptor (ER)1 mRNA, but not that of ER2, in MC3T3-E1 in a time-dependent manner. Total RNAs were isolated from MC3T3-E1 cells treated with bone morphogenetic protein (BMP)-2 or vehicle for the indicated periods, and estrogen receptor (ER)1 expression was examined by reverse transcription-polymerase chain reaction (RT-PCR) analysis. ER1 mRNA expression decreased from 6 to 24 hours after BMP-2 exposure (A). On the other hand, ER1 mRNA expression did not largely change over the period in the vehicle-treated MC3T3-E1 cells (B). Glyceraldehyde-3-phosphate dehydrogenase gene (GAPDH) expression was also analyzed as an internal control.
4. BMP-2 suppresses E2-induced ERE-mediated reporter activation in MC3T3-E1 cells
Next, the study observed whether the BMP-2-elicited downregulation of ER1 mRNA expression leads to reduced responsiveness of MC3T3-E1 cells to E2 by luciferase assay. As conveyed in Fig. 4, the E2-induced luciferase activity driven by the ERE-containing promoter was markedly suppressed when the cells were pretreated with BMP-2, indicating that BMP-2 desensitizes MC3T3-E1 osteoblastic cells to estrogen through the transcriptional downregulation of ER1.
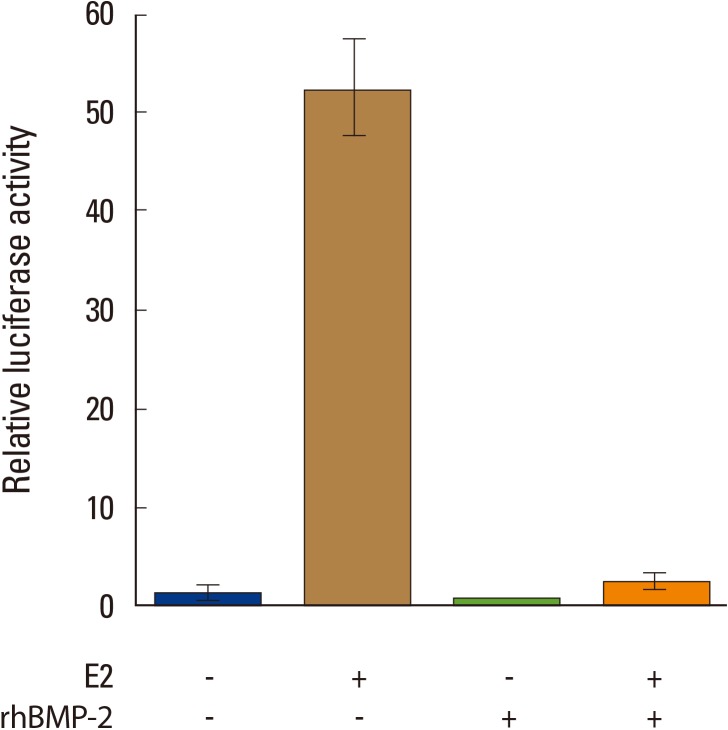
Bone morphogenetic protein-2 suppresses estradiol (E2)-induced estrogen responsive element (ERE)-mediated reporter activation in MC3T3-E1 cells. MC3T3-E1 cells transfected with pERE-G-luc, a reporter plasmid harboring the firefly luciferase gene under the control of a β-globin gene promoter, were pretreated with recombinant human bone morphogenetic protein-2 (rhBMP-2; 250 ng/mL) for 48 hours. These cells were then treated with estradiol (E2; 100 nM) for 24 hours, and the induced luciferase activities were evaluated. pCMVsport-β-gal, a reporter plasmid designed to express a β-galactosidase gene constitutively, was simultaneously transfected for normalization. The normalized values of the luciferase activity for the non-treated MC3T3-E1 cells are set at 1. Data are presented as the mean±SD of the three experiments. **P<0.01.
DISCUSSION
In the present study, a reporter system was established in order to identify the molecules that potentially regulate ER1 gene expression in mouse osteoblastic MC3T3-E1 cells. 7 molecules were evaluated in a pilot study. Further, it was demonstrated that rhBMP-2 among the molecules suppressed ER1 gene expression, leading to a restrained responsiveness of the cells to E2.
So far, a number of bioactive molecules, besides estrogens, including hormones, growth factors, cytokines and chemical compounds, have been shown to play a role in the regulation of osteoblast functions. More importantly, estrogens and these molecules have been reported to act on osteoblasts, not only independently, but also interactively.[11-15,17] However, information regarding bioactive molecules that modulate ER mRNA expression in estrogen-responsive cells remains limited. Of these molecules, EGF, 1,25-(OH)2D3 and its agonistic derivatives have been previously demonstrated to decrease the ER1 expression in MCF-7, an ER-positive human breast tumor-derived cell line.[18,19] Correspondingly, in this study, these two molecules decreased ER1 promoter activities in MC3T3-E1-MERAluc1 clones (Fig. 2), suggesting that the suppression of ER1 expression by these molecules could be a general event for many cell types. Conversely, estrogens have been reported to increase ER1 mRNA levels in MCF-7[20,21] as well as in the endothelium.[22,23] Against our expectations in the present study, however, the ER1 gene promoter activity was not affected by E2 and 4-OH-Tam, a tissue-selective agonist of ERs (Fig. 2). Considering that there have been no report so far that describes the presence of estrogen responsive elements (EREs) in the ER1 gene promoter, the discrepancy may be explained by assuming that the ER1 mRNA level may be post-transcriptionally regulated.
Very recently, Matsumoto et al.[24] have reported that estrogen exerts a stimulatory effect on osteoblast differentiation through the activation of signaling mediated by BMP-4, which is structurally similar to BMP-2. Corresponding to the results of this study, the authors also indicated that ER1 expression was significantly suppressed by BMP-4 treatment, suggesting the presence of a negative feedback loop for osteoblast differentiation.
Overall, the results presented herein indicate that the preferable effect of estrogens on bone metabolism may be abrogated by BMP treatment. Although it currently remains unknown whether our findings are true in vivo, this study will serve as a warning for the clinical use of BMPs. Further studies are currently ongoing in order to elucidate a mechanistic insight of our findings, and the results will be reported elsewhere.
ACKNOWLEDGEMENTS
The author thanks Dr. Hiroyuki Kawashima, a sometime professor at Niigata University Graduate School of Medical and Dental Sciences, for his helpful guidance and encouragement. This work was supported by Grant-in-Aid for Young Scientists (B) from the Ministry of Education, Culture, Sports, Science and Technology (MEXT) (No. 13771092) and by grants from Mochida Memorial Foundation for Medical and Pharmaceutical Research and Astellas Foundation for Research on Medicinal Resources.
Notes
No potential conflict of interest relevant to this article was reported.
Notes
This work was supported by a Grant-in-Aid for Young Scientists (B) from the Ministry of Education, Culture, Sports, Science and Technology (MEXT), and by grants from Mochida Memorial Foundation for Medical and Pharmaceutical Research and Astellas Foundation for Research on Medicinal Resources.